プレスリリース




卵を産む哺乳類カモノハシとハリモグラの苦味感覚を解明
(@Press) 2022年06月10日(金)11時00分配信

■ ポイント
●半水生のカモノハシと陸生のハリモグラは検知できる苦味物質の種類が大きく異なることを解明。
●ヒトを含む全ての哺乳類と同様に、植物などに含まれる有毒な配糖体を苦く感じることも解明。
●哺乳類進化の始まりである約2億年前から、植物毒を苦く感じる能力が維持されてきたことを示唆。
■ 概要
北海道大学大学院地球環境科学研究院の早川卓志助教と、明治大学研究・知財戦略機構の糸井川壮大研究推進員は、京都大学、アデレード大学、オーストラリア国立大学、コペンハーゲン大学との国際共同研究チームをつくり、卵を産む哺乳類(単孔類)であるカモノハシとハリモグラが持つ苦味センサータンパク質(苦味受容体)の機能を網羅的に分析しました。水中の多様な生物を食べるカモノハシは、幅広い種類の苦味物質を検知できる万能型苦味受容体を持つが、アリ・シロアリ食に特化したハリモグラは、カモノハシのような万能型受容体を持たず、検出できる苦味物質が少ないことが分かりました。
ハリモグラがアリやシロアリを専門に食べるようになったことで苦味の重要性が下がり、限られたものにしか苦味を感じなくなった一方で、水中で様々な生物を摂食するカモノハシは苦味受容体を使って食べられるものの選択を行っている可能性を示しています。
さらにカモノハシやハリモグラでも、植物などに含まれる有毒な配糖体を検知する苦味受容体は残されており、この苦味受容体の機能はヒトを含む全ての哺乳類グループで共通のものであることも分かりました。ヒトやその他の哺乳類がカモノハシ・ハリモグラと分かれたのは約2億年前まで遡ります。大型恐竜が繁栄し、花を咲かす被子植物の多様化が始まろうとしていた時代です。恐竜と競合しながら、植物や昆虫などの毒を含みうる食べ物を口にして進化した哺乳類において、苦味感覚の進化が非常に重要であったことを本研究は意味します。
なお、本研究成果は、日本時間2022年6月1日(水曜)公開のMolecular Biology and Evolution誌に掲載されました。
画像1: https://www.atpress.ne.jp/releases/313598/img_313598_1.png
研究対象の単孔類と有袋類。
単孔類(A)カモノハシ(B)ハリモグラ
有袋類(C)フクロネコ(D)タスマニアデビル(E)コアラ(F)タマーワラビー
■ 背景
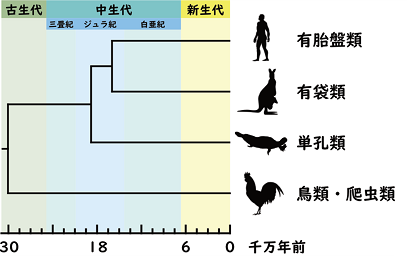
単孔類は私たちヒトと同じ哺乳類の仲間ですが、卵を産むという哺乳類の祖先が持っていた特徴を現在まで残す「生きた化石」とも呼べる存在です(上図、及び図1)。現在、単孔類はオセアニア地域に生息するカモノハシとハリモグラの2グループしかいませんが、それぞれが特徴的な生態を持っています。カモノハシは、湖沼や河川に生息し、水中で様々な無脊椎動物を食べて暮らしています。一方、ハリモグラは陸生で土や枯木の中のアリやシロアリを主食にしています。
私たちはものを食べる時、味や匂いを基に食べ物の良しあしを判断します。こうした味や匂いを検知するシステムは、環境に対して高い適応力があるため、生存に必須でなくなった感覚が退化したり、特殊な環境に適応するために新しい感覚が進化したりすることが様々な動物で観察されています。
研究グループは今回、毒物などの有害物質を検知し、適切な食物選択をするのに重要な味覚である苦味感覚に注目しました。脊椎動物の苦味感覚は、ゲノム中に数種類〜数十種類ある苦味受容体TAS2Rが苦味物質を受け取ることで発生します。苦味受容体の機能と食性の関係は、霊長類やコウモリの仲間ではよく研究されていますが、哺乳類の進化を明らかにするためには欠かせない、有袋類や単孔類の研究は、今までほとんどありませんでした。そこで本研究では、カモノハシとハリモグラはどんな苦味感覚を持っていて、食性とどう関係するのか、そして、胎生哺乳類(*1)の苦味受容体との比較によって、哺乳類の祖先はどんな苦味感覚を持っていたのか、という問いに挑戦しました(図1)。
■ 研究成果
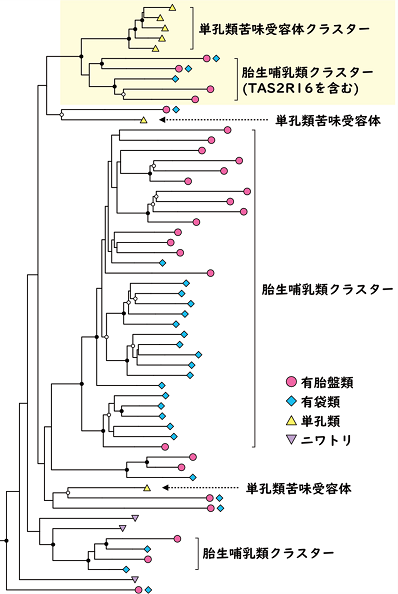
本研究の著者らを含む国際単孔類ゲノムプロジェクト(*2)において、単孔類は他の哺乳類と比べて苦味受容体遺伝子の個数が非常に少なく、ヒトは26種類持つのに対して、カモノハシは7種類、ハリモグラは3種類しか持っていないことが明らかにされていました。しかし、苦味受容体の中には単体で多様な苦味物質を受容できる「万能型受容体」が存在することも知られており、カモノハシやハリモグラが万能型受容体を持っているかどうかは不明でした。そこで研究グループはまず、この数少ない苦味受容体遺伝子が、非単孔類哺乳類(胎生哺乳類)のどの苦味受容体遺伝子と相同な関係にあるのかを調べました。
その結果、単孔類の苦味受容体遺伝子のほとんどが、有胎盤類の苦味受容体TAS2R16(*3)を含む遺伝子グループと同じグループに含まれており、染色体上でも相同な位置にあることが分かりました(図2)。TAS2R16は植物などに含まれる防御物質の一つであるβグルコシド(*4)と呼ばれる配糖体を検知する苦味受容体です。βグルコシドの中には分解されると有毒な青酸(シアン化水素)を発生させる青酸配糖体と呼ばれるものもあります(例:タピオカの原料であるキャッサバ芋のリナマリン、ユーカリの葉のプルナシン)。
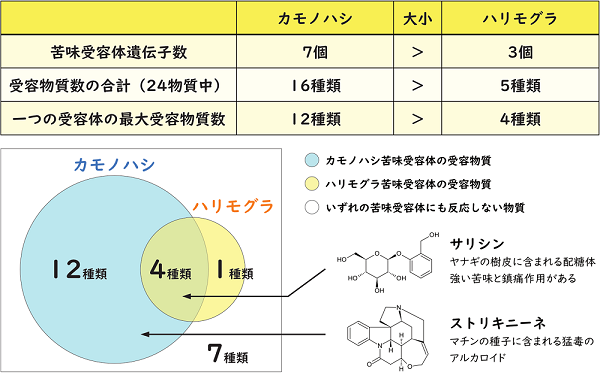
次に、カモノハシとハリモグラの苦味受容体がどんな物質に反応するのかを培養細胞を用いて解析しました(図3)。その結果、カモノハシは、使用した苦味物質24種類のうち18種類の物質を検知できました。また、驚くべきことに、使用した苦味物質の半数を一手に検知できる万能な苦味受容体を持っていることも判明しました。このことは、カモノハシが苦味受容体数から想定される以上に多様な苦味物質を検知できることを示しています。カモノハシは、微弱な電気や機械刺激を検知して濁った水の中で効率的に食べ物を探せますが、食べるか否かの判断には苦味感覚も利用していると考えられます。
一方、ハリモグラは、カモノハシが持つ万能型の苦味受容体を失っており、他の受容体もカモノハシよりも検知できる苦味物質の数が少ない傾向にありました。これは、ハリモグラがアリやシロアリに特化した食性を持っており、接触する苦味物質の種類が限られることが一因と考えられます。アリやシロアリといった限られたものしか食べないハリモグラにとって、多様な物質を検知できる苦味感覚はそれほど重要ではなく、限られた検知能力だけを残しているのかもしれません。
有胎盤類のTAS2R16と近縁な単孔類の苦味受容体の一部は、βグルコシドを受容することも分かりました。また、比較対象として使用した有胎盤類のTAS2R16と有袋類のTAS2R705(有胎盤類のTAS2R16と相同な有袋類の苦味受容体)がβグルコシドの受容体であることも分かりました。このことは、植物や一部の無脊椎動物が持つ防御物質であるβグルコシドを苦味として検知する能力が、現生の哺乳類全般に広く共有された重要な能力であることを意味すると同時に、この能力が単孔類と胎生哺乳類が分岐する以前の初期哺乳類で既に獲得されていたことを示唆します。単孔類と胎生哺乳類が分かれたのは恐竜が全盛の時代だった2億年近く前までさかのぼります。
しかし、現生植物に広く見られる有毒な青酸配糖体は、その生合成遺伝子の起源がさらに古い時代であるため、この当時には既に存在していたようです。つまり、有害な配糖体を苦味として検知する能力は、単孔類と胎生哺乳類が分岐する以前の初期哺乳類の採食選択に一定の役割を果たしていたことを示唆します。
■ 今後への期待
カモノハシやハリモグラは南半球の限られた地域にのみ生息し、野外での継続的な観察も難しいため、その生態には多くの謎が残されています。本研究では、培養細胞を用いた実験系を用いることで、苦味感覚という切り口から単孔類における遺伝子と生態の関連とその進化の一端を明らかにすることができました。しかし、今回解析した苦味受容体のうち、どの苦味受容体がカモノハシとハリモグラの舌や口の中で機能しているのかはまだ分かっていません。また、苦味感覚の生態的意義を明らかにする上で、野生下の個体が実際に食べているものに含まれる物質や接触する物質との関連を明らかにすることも重要です。
今後は、本研究を基礎として、舌や口の中の遺伝子発現の分析や苦味物質が採食行動に与える影響の分析によって、謎多き動物・単孔類の味覚機能とその生態的意義の詳細が明らかになることが期待されます。
■ 研究助成
本研究は、日本学術振興会・科学研究費助成事業(16K18630, 18J22288, 18H04005, 19K16241, 19K21586, 21KK0106, 21KK0130)、同・二国間交流事業『日本とオーストラリアの絶滅危惧哺乳類の地球縦断型比較ゲノム研究の確立』(JPJSBP 120219902)、日本科学協会・笹川科学研究助成『哺乳類の味覚の進化的起源の解明を目的としたオーストラリア産有袋類・単孔類の味覚受容体の分子生態学研究』(29-534)等の助成により実施されました。
■ 論文情報
論文名
Functional diversity and evolution of bitter taste receptors in egg-laying mammals(卵を産む哺乳類の苦味受容体の機能の多様化と進化)
著者名
Akihiro Itoigawa1,2*, Takashi Hayakawa3,4*, Yang Zhou5, Adrian D. Manning6, Guojie Zhang7, Frank Grutzner8, Hiroo Imai9*.
*責任著者(1明治大学農学部農芸化学科, 2京都大学霊長類研究所, 3北海道大学大学院地球環境科学研究院, 4日本モンキーセンター, 5BGI-Shenzhen, 6オーストラリア国立大学, 7コペンハーゲン大学, 8アデレード大学, 9京都大学ヒト行動進化研究センター)
雑誌名
Molecular Biology and Evolution(分子進化生物学の専門誌)
DOI
10.1093/molbev/msac107
公表日
2022年6月1日(水)(オンライン公開)
■ お問い合わせ先
北海道大学大学院地球環境科学研究院 助教 早川卓志(はやかわたかし)
TEL 011-706-4524 メール hayatak@ees.hokudai.ac.jp
URL https://noah.ees.hokudai.ac.jp/hayakawa/
明治大学研究・知財戦略機構/農学部農芸化学科 食品機能化学研究室
研究推進員 糸井川壮大(いといがわあきひろ)
メール ak.itoigawa0121@gmail.com
■ 配信元
北海道大学社会共創部広報課(〒060-0808 札幌市北区北8条西5丁目)
TEL 011-706-2610 FAX 011-706-2092
メール jp-press@general.hokudai.ac.jp
明治大学経営企画部広報課(〒101-8301 千代田区神田駿河台1-1)
TEL 03-3296-4082 FAX 03-3296-4087
メール koho@mics.meiji.ac.jp
■ 参考図
画像2: https://www.atpress.ne.jp/releases/313598/img_313598_2.png
図1. 主要な哺乳類の系統関係と分岐年代。ゲノム解析から単孔類と胎生哺乳類は1億8760万年前(ジュラ紀)に分岐したと推定されている。
画像3: https://www.atpress.ne.jp/releases/313598/img_313598_3.png
図2. 哺乳類の苦味受容体遺伝子の系統関係。単孔類の苦味受容体のほとんどがTAS2R16を含む胎生哺乳類苦味受容体クラスターと同じグループに含まれる(黄色部分)。
画像4: https://www.atpress.ne.jp/releases/313598/img_313598_4.png
図3. カモノハシとハリモグラの苦味受容体の機能差
■ 用語解説
*1)胎生哺乳類
ヒトのように発達した胎盤を持ち母親の子宮で胎児を育てる「有胎盤類」とカンガルーのように腹部にある袋(育児嚢)の中で未熟児を育てる「有袋類」の総称。
*2)国際単孔類ゲノムプロジェクト
カモノハシとハリモグラの全ゲノム配列を解読した国際プロジェクト。2021年に発表『カモノハシとハリモグラの全ゲノム解読に成功!〜世界でたった2グループしかいない「卵を産む哺乳類」のゲノムの進化を解明〜』
https://www.hokudai.ac.jp/news/2021/01/2-48.html
*3)TAS2R16
ヒトでよく研究されている苦味受容体の一つ。βグルコシドが主要な受容物質で、マンノースやゲンチオビオースなど他の糖に由来する配糖体も一部受容できる。
*4)βグルコシド
グルコースに由来する配糖体。植物に広く見られ、一部の昆虫も産生する。捕食に対する防御などに用いられる。例:サリシン(ヤナギの樹皮)、アルブチン(コケモモ)、リナリマリン(キャッサバ芋)、プルナシン(ユーカリの葉)
プレスリリース提供元:@Press
{kind=link}
{kind=link}
{kind=link}
{kind=link}